Mounir M Salem-Bekhit1,2 ![]() ,
Mohamed Osman Gad El Rab3,
Mahmoud M Tawfick1,
Mohammad Raish4,
Mohamed Dahmani Fathallah5,
Mohsen Bayomi3
,
Mohamed Osman Gad El Rab3,
Mahmoud M Tawfick1,
Mohammad Raish4,
Mohamed Dahmani Fathallah5,
Mohsen Bayomi3
For correspondence:- Mounir Salem-Bekhit Email: mbekhet@ksu.edu.sa Tel:+966501783166
Received: 7 November 2015 Accepted: 10 March 2016 Published: 30 April 2016
Citation: Salem-Bekhit MM, Gad El Rab MO, Tawfick MM, Raish M, Fathallah MD, Bayomi M. Humoral and cellular immune responses to modified hepatitis B plasmid DNA vaccine in mice. Trop J Pharm Res 2016; 15(4):687-693 doi: 10.4314/tjpr.v15i4.3
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To evaluate the immunogenicity and types of immune response of a quality-controlled modified recombinant hepatitis B surface antigen (HBsAg) plasmid encoding HBsAg in mice.
Methods: The characterized plasmid DNA was used in the immunization of Balb/c mice. Three groups of mice were intramuscularly injected with three different concentrations (50, 25 and 10 μg/100 μL) of the modified plasmid. Humoral immune response was monitored by enzyme-linked immunosorbent assay (ELISA), while cellular immune response was investigated by analysis of spleen cytokine profile (TNFα, IFN γ and IL2) as well as CD69 ex
Results: In general, the activated CD4 cells showing intracellular cytokines were higher than CD8 positive population of cells (p < 0.05). These findings indicate that the vaccine induced both a humoral and cellular immunity. Cytokine profile also showed high levels of TNFα, IFN γ and IL2 and CD69 ex
Conclusion: A 10 µg dose intramuscular injection of the modified DNA-based vaccine encoding HBsAg in mice induces both high humoral and cellular immune responses.
Introduction
Hepatitis B (HB) is a potentially life-threatening liver infection caused by Hepatitis B virus (HBV) [1]. It is an important worldwide health concern, more than 240 million are chronic carriers according to World Health Organization (WHO) [2]. DNA-based immunization is an efficient technique to induce humoral (antibody) and cellular (T cell) immune responses.
The use of DNA-based vaccines has recently attracted much attention and shown promising outcomes by stimulating the immune system [3]. The protection against HBV infection depends on strong antibody response. Th1-type and cytotoxic T cells (CTL) responses to HBV and the associated antiviral cytokines (TNF-α, IFN-α, and IL-2) might play a major role in virus resolution during natural infection.
Antigen presentation includes primarily foreign protein degradation in the host cells through association of proteolytic fragments with two different types of major histocompatibility complex (MHC) proteins (class I and class II MHC) which present degraded protein fragments to T-cells. Thus, the ability of DNA vaccine to elicit both humoral and cell mediated immune responses is a great advantage of using DNA vaccine [4].
The purpose of the present investigation was to assess the immunogenicity of previously reported DNA-HBsAg - based vaccine comprising modified gWiz-HBsAg plasmid, characterized and quality-controlled by Salem-Bekhit et al [5], and type of immune responses produced in mice.
Methods
Vector and expression plasmids
The gWiz™ expression vector, used as control in the quality control experiments, and the hepatitis B DNA vaccine recombinant plasmid gWiz™HBsAg (gWiz™ plasmid harbours HBsAg encoding nucleotide sequence) were purchased from Aldevron, USA. The full integrity and quality of the nucleotide sequence of gWizHBsAg plasmid was characterized and previously controlled previously [5]. The quality controlled recombinant plasmid gWizHBsAg was used in mice immunization experiments performed in this study.
Modified gWiz™HBsAg plasmid mice immunization
The recombinant modified gWizHBsAg plasmid (6.435 kbp long and had a pUC origin of replication. Aldevron, USA). previously reported by Salem-Bekhit et al [5]. Briefly, The gWi zHBs have sequentially propagated in Escherichia coli DH5α strain, Genotype: F- endA1 glnV44 thi-1 recA1 relA1 gyrA96 deoR nupG Φ80dlacZΔM15 Δ(lacZYA-argF)U169, hsdR17(rK- mK+), λ. The gWiz high-Expression HBsAg plasmid (gWizHBsAg) 5 mg. The prepared plasmid was analyzed, using agarose gel electrophoresis, DNA restriction profiling and DNA sequencing. The gWizHBsAg plasmid was used for immunization of 21 female Balb/c mice aged 6–8 weeks bred in King Saud University (KSU) animal house and kept under specific pathogen free conditions. These mice were divided into three groups (A, B and C), each group comprised seven mice. Mice received intramuscular (IM) injections by insulin syringe (Becton-Dickinson, Franklin Lakes, NJ, USA) in the quadriceps muscles of the hind limbs. The three groups A, B and C received different doses of gWizHBsAg plasmid DNA dissolved in 100 μL of normal saline 50 μg/100 μL, 25 μg/100 μL and 10 μg/100 μL, respectively. Mice were boosted with a dose of 4 μg after 2 weeks from the initial immunization. Ethical approval for the animal study was obtained from the Ethics Committee of the College of Pharmacy, KSU, Riyadh, Saudi Arabia.
Mice bleeding and detection of specific antibodies against HBsAg
Serial bleedings were performed at different time points starting with pre-immunization bleeding at the beginning and then at weekly interval up to 6 weeks after the first immunization. The blood samples were analyzed for the presence of HBsAg-specific IgM antibodies from week 0, 1, 2, 3 and 4 and starting from weeks 3, 4, 5 and 6 for the detection of HBsAg-specific IgG antibodies. Microtitre plates of ELISA kits (Alpha Diagnostic International, U.S.A) coated with HBsAg protein were used for the detection anti-HBsAg antibodies. In brief, 100 μL of serum diluted in 1:200 was added to each well then incubated for one hour at 25 oC. After the samples were washed with PBS, bound proteins were detected with HRP-conjugated goat anti-mouse IgM or anti-mouse IgG. Color was generated by adding reaction stop solution and the absorbance was measured at 450 nm by ELISA reader. ELISAs were performed and HBsAg specific IgM and IgG antibodies were calculated according to the manufacturer’s instructions.
Isolation and culturing of splenocytes isolated from immunized mice
To determine the cellular reaction of the modified gWizHBsAg plasmid immunization, the spleen, following six-week immunization course, were isolated from the three groups of mice, A, B, and C, and splenocytes were cultured as follows. Briefly, immediately after mouse sacrifice, spleen was put in a clean Petri-dish with a suitable tissue culture medium, disrupted mechanically using a 5 ml syringe head after cutting into small pieces and passed through a nylon cell strainer (Becton Dickinson, New Jersey, USA). Contaminating red blood cells were lysed using 0.83 % ammonium chloride and incubated at 37 oC for 5 min. In ice-cold RPMI 1640 tissue culture medium supplemented with 10 mM fetal bovine serum, 10 mM HEPES, 2 mM glutamine, 100 U/mL streptomycin and 100 U/mL penicillin the cells were suspended.
Finding of splenocytes surface markers and intracellular cytokines
HBsAg-specific CTL assay was performed using panels of monoclonal antibodies used for the detection of surface markers and intracellular cytokines of immunized splenocytes. Splenocyte concentration of 2 x 106 /mL was stained in 4 test tubes:- 1- CD4 FITC, CD69 PE and Tumor Necrosis Factor (TNF) precp, 2- CD4 FITC, IL2 PE and INF-γ precp., 3- CD8 FITC, CD69 PE and TNF precp., 4- CD8 FITC, Interleukin-2 (IL2) PE and INF-γ precp. Surface marker monoclonal antibodies were added first to stain the cells for 20 min in the dark, cells were then washed with cold 5 % BSA/PBS and the pellet was re-suspended in complete RPMI. The cells were fixed and permeablized with 100 μL of BD Cytofix/Cytoperm buffer and incubated for 20 min at room temperature. Thereafter, 20 μL of cytokine monoclonal antibodies were added, incubated for 15 min, washed 3 times and suspended in BD Pharmingen stain buffer. Data acquisition and analysis were obtained by flow cytometer.
Statistical analysis
The data were processed by SPSS 11.5 software. For each set of data, mean and standard deviation (SD) were calculated. IgM and IgG values were plotted against time and histograms were built to compare the values obtained for cytokines. Statistical significance of differences was defined as p < 0.05.
Results
DNA plasmid encoding HBsAg induces humoral immune response in BALB/c mice
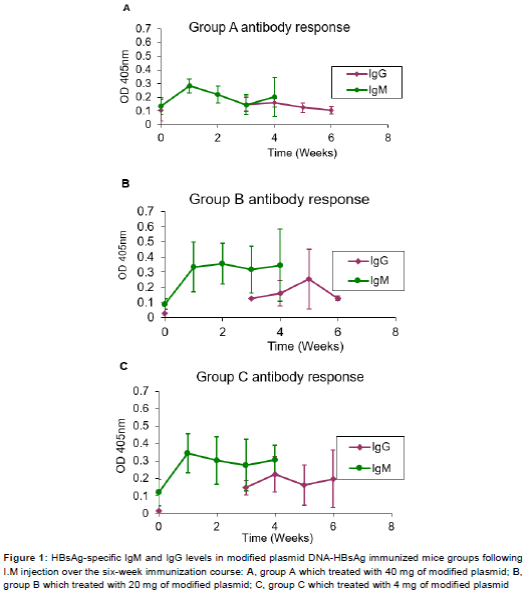
The humoral immunity was evaluated by detecting specific HBsAg IgG and IgM induced by modified DNA HBsAg plasmids in BALB/c immunized mice. ELISAs results showed that HBsAg specific IgG and IgM antibodies were detectable in the sera of the immunized mice groups A, B and C. The baseline reading of the pr-immunization bleed sera showed an OD450nm of 0.15. The group A mice, which received a total of 20 mg of modified plasmid DNA-HBsAg over the six-week immunization course, demonstrated IgM antibodies as early as one week after the first immunization showing a detectable signal at an OD450nm of 2.8. However, in the following 2nd and 3rd weeks showed a gradual drop to an OD450nm of 1.5. However, at the 4th week, the IgM level showed a rise up to OD450nm of 2.0. IgG antibodies were detectable starting from the 3rd week after the first immunization giving an OD450nm of 0.18. Thereafter, IgG level started to decrease gradually in the 5th and 6th weeks to 0.1 OD450nm. The collective results for IgM and IgG levels for group A mice are shown in A.
In immunized mice group B, which received a total of 10 mg of modified DNA-HBsAg plasmid by the I.M. route, IgM antibodies were detectable at a level of 0.35 OD450nm in the sera of the first bleed sample after one week of immunization. In the following bleedings at the 2nd, 3rd and 4th week, the IgM antibodies levels were almost maintained. On the other hand, IgG antibodies were detectable at a level of 1.0 OD450nm in the 3rd week. The level of IgG antibodies was then increased to 0.15 OD450nm in the 4th week and to 2.5 OD450nm in the 5th week. Afterwards, the level of IgG antibodies started to decrease gradually to a level of 0.1 OD450nm at the 6th week. The collective results for IgM and IgG levels for this group are shown in B.
In group C mice which received a total of 4 mg of modified plasmid DNA-HBsAg over the immunization course, IgM antibodies level were detectable after one week from the first immunization of 0.35 OD450nm. The level dropped gradually to 0.3 and 0.29 OD450nm in the 2nd and 3rd week bleeds, respectively, and then increased to 0.3 OD450nm at the 4th week. The IgG antibodies level were detectable in a level of 0.15 OD450nm at the 3rd week, then the level increased to 0.22 OD450nm in the 4th week and dropped to 0.15 OD450nm at 5th week. At the 6th, week the IgG antibodies level again increased to 0.2 OD450nm. The collective IgM and IgG antibodies levels for this group are shown in C.
Activation of cellular immunity in BALB/c mice
The cellular immunity in DNA-HBsAg plasmid immunized BALB/c mice was assessed by detecting the CD4+ and CD8+ cells. Activation of CD4+ and CD8+ cells was examined in the three groups of immunized mice by measuring the number of cells showing intracellular cytokine release, TNF, INF-γ and IL-2. The activation marker CD69 was also measured.
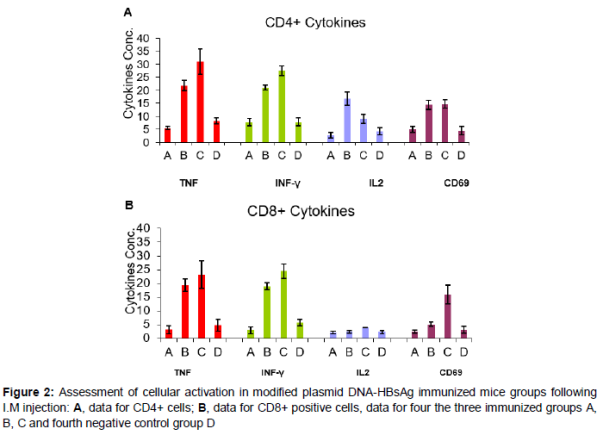
Activation of CD4+ cells for the three groups of immunized mice is shown in A. It was found that 30 % of CD4+ cells in group C mice were actually producing TNF as compared to 21 and 5 % in groups A and B, respectively. For INF-γ as shown in A, 28 % of CD4+ cells from mice group C showed detectable intracellular INF-γ. While, in groups B and A, mice 20 and 8 % of the CD4+ cells were actively producing INF-γ, respectively. CD4+ cells from group B mice showed 18 % of activated cells secreting IL-2, while, it is 10 % in group C and only 2 % in group A. The activation marker, CD69, was detected in equal number in both group B and group C where 14 % of the CD4+ cells were positive, however, in group A, only 4 % of the CD4+ cells were positive for the marker.
In the case of CD8+ cells (B), intracellular TNF was detected in 23 % of cells in group C, mice while 18 % in group B and 2.5 % in group A were secreting TNF. For INF-γ, CD8+ cells from group C mice showed also the highest number of activated cells secreting INF-γ (24 %). In group B, mice 18 % of cells were active and in group A only 4 % of cells showed INF-γ. IL-2 was detected in 5 % of the cells from mice group C; mice while in groups A and B, only 2 % were positive for IL-2. CD8+ cells from group C mice showed 15 % of cells expressing CD69, while the marker was only detected in 2 and 5 % of CD8+ cells in groups A and B, respectively.
Discussion
The best way to prevent hepatitis B is by receiving hepatitis B vaccine. It is important to accelerate the vaccination schedule against hepatitis B because it might increase patient compliance in addition to offering earlier protection for the high-risk group or environment.
HBV infection is considered to be one of the dominant problem in most countries worldwide, specifically those in the Middle East, Asia, and Africa [6-8]. Hepatitis B infection is the major cause of diseases of liver such as cirrhosis and hepatic carcinoma. Despite the occurrence of HBV infection that has declined fallowing mass HB vaccination programs, the average incidence of chronic HBV infection internationally is estimated at 6.6 % [7]. The rate of HbsAg positivity among the vaccinated group was 0.8 % in comparison with 2.2 % among the non-vaccinated group. The study concluded that the efficacy of HB vaccine in inhibiting the carriage of HbsAg, 5 years after full course vaccination, was assessed to be around 67 % in Egypt [9].
One month [10] after the third dose, the seroprotection was (96.5 %) with mean geometric anti-HBs titers of 2 560.0 mIU/mL was indicated through the experimental data that were obtained during the trial course. In addition, twenty subjects as a whole in the age group of 40-49 years (11/307; 3.5 %) had resulted an antibody titer of 8.60 mIU/mL and presented a low response to the vaccine. While all tested subjects attained 100 % seroconversion [11], risk factors associated with non-response to hepatitis B vaccine included age, gender, history of smoking and route of administration of vaccine [12].
Viral vectors and recombinant DNA have been used for years in human clinical trials and in laboratory animals as vectors for Ag(s) of interest to make strong effector and memory CD4_ and CD8_ T cell responses in both the treatment and inhibition of diseases [13-15].
Results from this study show that both humoral and cellular immune responses were induced in mice by the intramuscular injection of a modified plasmid DNA-HBs Antigen. In all three groups of mice, A, B and C which received 20, 10 and 4 µg of vaccine, respectively, the antibodies of IgM class were detectable at a comparatively high concentration in the 3rd week.
This finding confirms the induction of a primary response to the injected vaccine (16). It has been documented that specific cellular immunity against HBV is a key factor in the control of HBV infection [17,18]. IgM level decreased gradually in all three groups over the following two weeks and then showed a rise at the 4th week. As a consequence of the booster doses given to the animals, IgG type of antibodies appeared the 3rd week in all the groups. This indicates a normal secondary response to the injected vaccine. The IgG level started to decrease gradually thereafter. It is also noted that IgM showed higher levels as compared to IgG. This could be due to the concentration of the booster doses administered.
Compared to groups A and B, group C showed the best results. This may be taken to indicate that the best immunizing dose in this experiment lies in the range of 4 µg. In this group, higher levels of IgM were detected and showed a sustained level up to the 4th week. IgG antibodies became detectable at the 3rd week and also maintained an almost constant level up to the 6th week. Cellular responses to the modified DNA-HBs antigen vaccine were assessed in splenocytes isolated from the immunized mice by measuring intracellular cytokines (IFN-γ, TNF, IL-2 and the activation marker CD69) in CD4+ and CD8+ cells. In general, the percent of activated CD4+ cells showing intracellular cytokines was higher than the CD8+ population of cells. This is expected since CD4+ helper cells are usually higher than CD8+ cytotoxic cells.
Another observation worthy of note, the cytokines detected IFN-γ and TNF indicates a Th2 type of response. Viral antigens are known to drive T-helper cells into the Th2 type. It is also interesting to note that group C mice again showed the best cellular response. As appeared in the results, IL-2 was the least cytokine detected and the number of CD8+ cells secreting this cytokine was very low. In summary, results from this experimental study in BALB/c mice demonstrated that I.M. injection of a DNA-HBs antigen in a dose of 10 µg induced both humoral and cellular immune responses. Further studies may be needed to adjust the dose and timing of the booster doses to give a sustained immune response. Engaging therapeutic hepatitis B vaccines is a new approach against hepatitis B infection [19,20]. The cytokines (IFN-γ, TNFα, IL-2 and CD69) in CD4+ and CD8+ cells perform a vital role in various stages of viral infection [21-24]. However, experiments which are carried out in vivo and in vitro showed that secretion of Th1 cytokines (IFN-γ and TNF-α) play an important part in HBV elimination from hepatocytes [25,26]. Hepatitis B vaccine administered through gene therapy showed enhanced cytokines level in patients with Hepatitis B. The direct administration of cytokines seems to be a more efficient method for viral infection removal [20].
Conclusion
The findings of this study demonstrate that I.M. injection of a DNA-HBs antigen at a dose of 10 µg induces both humoral and cellular immune responses in BALB/c mice. Further studies are required to adjust the dose and timing of the booster doses to give sustained immune response.
Declarations
Acknowledgement
References
Archives


News Updates